More than One Way to the Egg
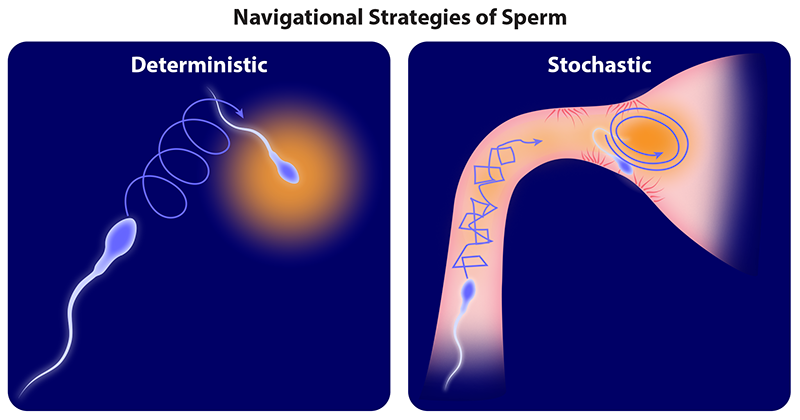
For all of us, fertilization was that singular and momentous occasion in which two gametes fused to trigger an irreversible chain of events that led to the formation of a new life. This form of reproduction is critical for driving genetic diversity and has been retained across the eukaryotic tree of life, from single-celled organisms to animals to plants. For well over 100 years, scientists have puzzled over the precise sequence of events by which gametes locate each other across a complex and heterogeneous landscape. The canonical view is that sperm steer themselves deterministically toward their target, the egg. Now, in a new study, Meisam Zaferani and Alireza Abbaspourrad of Cornell University show that bull sperm employ a distinctive biphasic (or two-part) swimming strategy in response to chemotactic cues, which can be further modulated by the surrounding fluid’s rheological properties [1]. This finding raises an intriguing possibility: sperm navigate the intricate confines of the female reproductive tract stochastically.
In the race for life, we picture a sperm undulating its flagella to swim resolutely toward a stationary egg. This image of the protagonists, though compelling, is not necessarily accurate. For one thing, in many single-celled organisms like algae and ciliates, both sexual partners are motile. In this case, the challenge of fertilization is a physically daunting one of trying to strike a moving target amid a cacophony of background noise [2].
But what is the mechanism of this search and interception process? How does it differ between species? Is it deterministic or stochastic? In nature, fertilization can take place either internally or externally (Fig. 1). The sperm of marine invertebrates, for external fertilization in open water, must somehow locate a remote egg, relying on any weak biochemical cues released by the egg to guide them along the way [3]. In contrast, in mammals, where fertilization is internalized, sperm cells also respond to chemical cues. But they must also adjust the beat of their flagella to chart a tortuous route through the microarchitecture of the female reproductive tract to the oviduct, often under intense competitive selection [4].
A navigating sperm cell will therefore experience distinct physical and molecular interactions, depending on the context and the desired type of fertilization. The precise functional relationship between external cues and the intrinsic flagellar beat, and the resulting swimming trajectory, has not yet been elucidated except in a few select species, and even then, only for some of the molecules that are involved in sensing and responding to signals [3]. Certain physical properties of the fluid environment, including viscosity, viscoelasticity, and even ionic composition, can also significantly impact the beat pattern [5].
Zaferani and Abbaspourrad used a controlled microfluidic environment and phase-contrast microscopy to monitor the response of bull sperm in two viscosity regimes to a potent potassium-channel blocker, 4AP. The drug is thought to mimic the biochemical cues released by the egg. In a viscoelastic buffer representative of mammalian bodily fluids, the researchers discovered that as 4AP concentration increased, the sperm swam in increasingly tighter circles. The researchers termed this behavior “chiral.” But in a low-viscosity buffer with no 4AP, the sperm progressed along linear paths, and their movement had a three-dimensional component that was absent in the viscoelastic case. And when the 4AP concentration increased, the sperm became more energetic and lost directionality. The researchers termed this behavior hyperactive. In both regimes, 4AP likely acted by inducing changes in the flagellar dynamics and tuning the asymmetry of the beat pattern.
The researchers developed a model to account for key features of the sperm’s chiral and hyperactive swimming, including speed, rolling, and flagellar asymmetry. They then used the model to derive statistical characteristics of the trajectories, such as orientational persistence and diffusivity, from their observations of the sperm. Intriguingly, which motility pattern dominated—chiral or hyperactive—depended strongly on the fluid’s rheological properties. Also, for both phenotypes, effective diffusivity decreased with increasing 4AP concentration. The apparent suppression of three-dimensional rolling in the non-Newtonian regime may be analogous to the wobbling-to-swimming transition recently identified in bacteria swimming in viscoelastic media [6]. Zaferani and Abbaspourrad concluded that a combination of biochemical and rheological cues can tune the biphasic motility strategy of mammalian sperm by adjusting the scrambling rate of the swimming trajectory and by modulating path diffusivity. They hypothesized that in a spatially heterogeneous environment a stochastic search may be more effective than a deterministic one in which the helical trajectory of the cell aligns gradually to a stimulus from a fixed, remote source [7].
When and how did the navigation strategies of sperm of different species diversify? In the fossil record, the first evidence of sexual reproduction in early eukaryotes (single-celled organisms with a nucleus) appeared over 1 billion years ago. Stochastic search strategies based on temporal comparisons, such as run-and-tumble chemotaxis, are typically associated with small prokaryotes (single-celled organisms without a nucleus), whose tiny bodies are overwhelmed by thermal noise. Larger eukaryotes access deterministic taxes [8]. But there are exceptions. For example, eukaryotes very closely related to the members of the animal kingdom, the choanoflagellates, adopt a stochastic search strategy to find higher concentrations of dissolved oxygen [9]. Zaferani and Abbaspourrad’s work highlights the need for more comparative studies that span the eukaryotic tree of life.
Fertilization is not all about the sperm though. The oviduct is lined with cells whose cilia (hair-like structures) coordinate dynamic background flows to guide and select sperm [10]. The response of sperm to viscosity gradients and mechanical contact may further interface with chemokinetic responses. It is likely that the hyperactive mode that involves sharp reorientations can facilitate sperm escape from physical boundaries. Thus, the stochastic chemokinetic behaviors of sperm and other small eukaryotes constitute a hitherto underappreciated search strategy whose theoretical and biomolecular underpinnings await to be fully explored.
References
- M. Zaferani and A. Abbaspourrad, “Biphasic chemokinesis of mammalian sperm,” Phys. Rev. Lett. 130, 248401 (2023).
- J. Pan and W. J. Snell, “Signal transduction during fertilization in the unicellular green alga, Chlamydomonas,” Curr. Opin. Microbiol. 3, 596 (2000).
- J. F. Jikeli et al., “Sperm navigation along helical paths in 3D chemoattractant landscapes,” Nat. Commun. 6, 7985 (2015).
- S. S. Suarez, “Mammalian sperm interactions with the female reproductive tract,” Cell Tissue Res. 363, 185 (2015).
- J. S. Guasto et al., “Flagellar kinematics reveals the role of environment in shaping sperm motility,” J. R. Soc., Interface 17, 20200525 (2020).
- S. Kamdar et al., “The colloidal nature of complex fluids enhances bacterial motility,” Nature 603, 819 (2022).
- B. M. Friedrich and F. Jülicher, “Steering chiral swimmers along noisy helical paths,” Phys. Rev. Lett. 103, 068102 (2009).
- K. Y. Wan and G. Jékely, “Origins of eukaryotic excitability,” Philos. Trans. R. Soc., B 376, 20190758 (2021).
- J. B. Kirkegaard et al., “Aerotaxis in the closest relatives of animals,” eLife 5 (2016).
- S. Li and W. Winuthayanon, “Oviduct: roles in fertilization and early embryo development,” J. Endocrinol. 232, R1 (2017).